РНК-полимераза - фермент, осуществляющий синтез молекул РНК. В узком смысле, РНК-полимеразой обычно называют ДНК-зависимые РНК-полимеразы, осуществляющие синтез молекул РНК на матрице ДНК, то есть осуществляющие транскрипцию. Ферменты класса РНК-полимераз очень важны для функционирования клетки, поэтому они имеются во всех организмах и во многих вирусах. Химически РНК-полимеразы являются нуклеотидил-трансферазами, полимеризующими рибонуклеотиды на 3"-конце цепи РНК.
Промотор - последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как стартовая площадка для начала специфической или осмысленной транскрипции. У прокариот промотор включает ряд мотивов, важных для узнавания его РНК-полимеразой, в частности так называемые последовательности -10 и -35. Промотор асимметричен, что позволяет РНК-полимеразе начать транскрипцию в правильном направлении и указывает то, какая из двух цепей ДНК будет служить матрицей для синтеза РНК.
Промоторный участок в пределах оперона может частично перекрываться или вовсе не перекрываться с операторным участком цистрона (гена).
То, под каким промотором находится кодирующий РНК участок ДНК, играет решающее значение в интенсивности экспрессии этого гена в каждом конкретном типе клеток. Активация промотора определяется присутствием в каждом типе клеток своего набора транскрипционных факторов.
Терминатор - последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как сигнал к прекращению синтеза молекулы РНК и диссоциации транскрипционного комплекса.
Часто эти последовательности заканчиваются цепочкой тиминовых нуклеотидов (у транскриптов - уридиновых), которой предшествует участок, содержащий внутренние, взаимно комплементарные последовательности в противоположных ориентациях («шпилечные» структуры). Область «шпильки» обогащена GC парами, придающими этой структуре большую устойчивость.
78. Отличие процесса транскрипции в разных клетках. Мозаичность строения генов. Синтез гетероядерной РНК, преобразование ее в информационную РНК. Сплайсинг. Процессинг.
Некоторые вирусы (такие как ВИЧ, вызывающий СПИД), имеют возможность транскрибировать РНК в ДНК. ВИЧ имеет РНК-геном, который встраивается в ДНК. В результате, ДНК вируса может быть объединено с геномом клетки-хозяина. Главный фермент, ответственный за синтез ДНК из РНК, называется ревертазой. Одной из функций ревертазы является создание комплементарной ДНК (кДНК) из вирусного генома. Ассоциированый фермент рибонуклеаза H расщепляет РНК, а ревертаза синтезирует кДНК из двойной спирали ДНК. кДНК интегрируется в геном клетки-хозяина с помощью интегразы. Результатом является синтез вирусных протеинов клеткой-хозяином, которые образуют новые вирусы. В случае с ВИЧ так же программируется апоптоз (смерть клетки) Т-лимфоцитов. В иных случаях клетка может остаться распростанителем вирусов.
Некоторые клетки эукариотов содержат фермент теломеразу, так же проявляющую активность обратной транскрипции. С её помощью синтезируются повторяющиеся последовательности в ДНК. Теломераза часто активирутся в раковых клетках для бесконечной дупликации генома без потери кодирующей протеины последовательности ДНК.
Отличительная черта строения многих генов эукариот - мозаичность (прерывистость) структуры смысловой части генов.
Это отличие связано с чередованием смысловых участков, несущих информацию о последовательности аминокислот в белке - экзонов, и участков некодирующих последовательностей, которые по длине значительно больше экзонов - интронов. В результате общая длина гена оказывается больше, чем можно было ожидать, примерно, в 5-7 раз.
В начале гена (до его смысловой части) находятся участки, которые обеспечивают правильную регуляцию работы гена. Их делят на две группы: неспецифические (одинаковые для всех генов) и специфические (характерные только для данного гена). Неспецифические регуляторные участки у эукариот называют «TATA-БОКС». В них чередуются тимин и аденин. Этот участок лежит на 30 нуклеотидов левее начала считывания гена. Установлено, что РНК-полимераза так ложится на ДНК, что ее опознающая часть закрывает TATA-БОКС, а активный центр РНК-полимеразы оказывается над первым считываемым нуклеотидом. Далее следует промоторный участок, состоящий из сайта-узнавания, сайта-связывания и сайта-инициации (А или Г) транскрипции. Комбинация нуклеотидов в промоторе такова, что если рамка считывания устанавливается неверно, то получается последовательность, характерная для стоп-кодонов, и транскрипция останавливается. За промоторным участком следует палиндром («перевертыш»), или инвертированный повтор. Это двухцепочечная последовательность ДНК, которая одинаково читается в обоих направлениях («казак» - справа налево и слева направо читается одинаково). Иногда такую последовательность называют «областью с двойной симметрией», где осью симметрии является центральная точка, относительно которой последовательность остается одинаковой. Важное свойство палиндромов - возможность образовывать шпильки в РНК или структуры креста - в ДНК. Если рамка считывания установлена неверно, то палиндром ДНК превращается в «крест», что делает невозможным дальнейшее продвижение фермента. Иногда палиндромы могут находиться в конце гена после его смысловой части и также, образуя «крест», будут прекращать синтез мРНК. Смысловая часть гена, как мы уже писали выше, состоит из чередующихся интронно-экзонных последовательностей. В структурной части гена расположены промотор и палиндром.
В последнее время выделяют включения специфической регуляции гена - энхансеры. Они расположены впереди гена на расстоянии в сотни и тысячи нуклеотидных пар. У эукариот существуют специальные регуляторные белки, опознающие энхансер и присоединяющиеся к нему. Таким образом происходит активизация гена. Геном образован не только ядерными, но и цитоплазматическими генами. В геноме гены выполняют разные функции. В зависимости от выполняемых функций гены подразделяют на обязательные гены, они кодируют жизненно важные функции в клетках всех типов (иногда их называют генами «домашнего хозяйства»), структурные гены, регуляторные гены, пунктуационные гены. В то время, как мРНК прокариот (бактерий и архей), за редкими исключениями, сразу готовы к трансляции и не требуют специальной обработки, эукариотические пре-мРНК подвергаются интенсивным модификациям. Так, одновременно с транскрипцией на уже синтезированном участке мРНК происходит «редактирование»(сплайсинг). В процессе сплайсинга из пре-мРНК удаляются не кодирующие белок последовательности - интроны, на 5" конец молекулы добавляется специальный модифицированный нуклеотид (кэп), на 3" конец добавляются несколько аденинов, так называемый полиадениновый хвост. Кэп узнаётся факторами инициации, белками, отвечающими за присоединение к мРНК рибосомы, полиадениновый хвост связывается с со специальным белком, ПАБ. Обычно эти посттранскрипционные изменения мРНК эукариот обозначают термином «процессинг мРНК». Полиаденилирование необходимо для транспорта большинства мРНК в цитоплазму и защищает молекулы мРНК от быстрой деградации (увеличивает время их полужизни). Лишенные поли-А участка молекулы мРНК (например, вирусные) быстро разрушаются в цитоплазме клеток эукариот рибонуклеазами.
Сплайсинг (от англ. splice - сращивать или склеивать концы чего-либо) - процесс вырезания определенных нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга РНК. Наиболее часто этот процесс встречается при созревании информационной РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки - экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки. Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК) и других некодирующих РНК.
Процессинг РНК
Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. К 5΄-концу присоединяется кэп, а к 3΄-концу поли-А хвост, который увеличивает длительность жизни иРНК. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемым единой последовательностью нуклеотидов ДНК, - альтернативный сплайсинг.
Узнаваемая РНК-полимеразой как стартовая площадка для начала специфической, или осмысленной, транскрипции . У прокариот промотор включает ряд мотивов, важных для узнавания его РНК-полимеразой, в частности так называемые последовательности -10 и -35. Промотор асимметричен, что позволяет РНК-полимеразе начать транскрипцию в правильном направлении и указывает то, какая из двух цепей ДНК будет служить матрицей для синтеза РНК. Промоторный участок в пределах оперона может частично перекрываться или вовсе не перекрываться с операторным участком цистрона (гена).
Wikimedia Foundation . 2010 .
Смотреть что такое "Промотор (молекулярная генетика)" в других словарях:
Эта страница глоссарий. См. также: Список генетических пороков развития и заболеваний Термины генетики в алфавитном поряд … Википедия
Нокаутные мыши Генетическая инженерия (генная инженерия) совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие… … Википедия
Группа функционально связанных между собой Генов, детерминирующих синтез белков ферментов, относящихся к последовательным этапам какого либо биохимического процесса. Концепция О. как часть теории генетической организации и регуляции… …
I Транскрипция (от лат. transcriptio переписывание) письменное воспроизведение слов и текстов с учётом их произношения средствами определённой графической системы. Т. бывает научная и практическая. Научная Т. применяется в лингвистических … Большая советская энциклопедия
Транскрипция в биологии, осуществляющийся в живых клетках биосинтез рибонуклеиновой кислоты (РНК) на матрице ‒ дезоксирибонуклеиновой кислоте (ДНК). Т. ‒ один из фундаментальных биологических процессов, первый этап реализации генетической… … Большая советская энциклопедия
Промотор-регуляторная последовательность 5’ области гена, определяющая место прикрепления РНК-полимеразы к ДНК. В промоторе находятся две последовательности, играющие важную роль инициации ТАТА-бокс и домен ЦЦААТ. Промотор указывает РНК-полимеразе с какого места начать синтез.
102вопрос. Поясните понятия: транскрипт и транскриптом.
Транскрипт –молекула РНК,образующаяся в результате транскрипции участка ДНК
Транскриптон -совокупность всех транскриптов синтезируемых одной клеткой, включает мРНК и некодирующую РНК
Вопрос.Какие молекулы синтезирует РНК-полимераза I.
Синтезирует 5,8sрРНК, 18sрРНК, 28sрРНК
отвечает за синтез крупныхрРНК, она локализована в ядрышке.
Вопрос.Какие молекулы синтезирует РНК-полимераза II.
Синтезирует мРНК, мяРНК(малоядерные), мкРНК, миРНК(малоинтерферирующие),
иРНК, она локализована в цитоплазме
Вопрос.Какие молекулы синтезирует РНК-полимераза III.
Синтезирует 5sРНК, тРНК, часть мяРНК, мелкиерРНК
Вопрос.К чему сводится механизм транскрипции (синтез РНК на матрице ДНК).
В результате транскрипции образуется первичныйтранскриптиРНК.
107вопрос.Что получило название – процессинг РНК. Назвать из каких отдельных процессов он складывается.
Процесс созревания иРНК называется – процессингом. Кэпирование, полиаденилирование, сплайсинг
Вопрос.Что представляет сплайсинг-реакция РНК-транскриптов. Роль сплайсингосомы в этом процессе.
Сплайсинг – это вырезание интронов из молекулы иРНК и сшивание экзонов при помощи ферментов лигаз.
Сложившийся макромолекулярный процесс в нем принимает участие большое количество макромолекул. Макромолекулярная структура осуществляется(сплайсингосома->малоядерные РНК). мяРНК распознают границу экзонов и интронов и связываются с ними.
109вопрос.Когда происходит и к чему сводится кэпирование 5′-конца РНК у эукариот. Транскрипты каких РНК-полимераз подвергаются кэпированию.
Сразу после транскрипции. К 5" концу иРНК присоединяется метилированный остаток гуанозина, эта структура называется кэп (шапочка). Кэп способствует связыванию иРНК с рибосомой в цитоплазме.
Только транскриптыРНК-полимеразыII
Вопрос.Перечислить все положения значения кэпированиямРНК.
Обеспечивает эффективность дальнейшей транскрипции.
Защищает транскрипт от деградации 5’-экзонуклеазами(5’-5’ связи)
Способ дальнейшему ходу процесса: стимулирует 3’-полиаденилирование и сплайсинг.
Требуется для экспорта мРНК из ядра
Обеспечивает связывание мРНК с рибосомой в цитоплазме
111.Что представляет собой полиаденилирование 3′-конца мРНК. Какую роль играет этот процесс. Транскрипты каких РНК-полимераз подвергаются этому процессу.
Полиаденилирование – к 3" концу молекулы иРНК присоединяется от 100 до 200 адениловых нуклеотидов. Образуется поли-А-участок. Этот участок стабилизирует молекулу иРНК, и способствует ее выходу из ядра в цитоплазму.
Транскрипты РНК -полимеразы II
Что представляет собой альтернативныйсплайсинг. Какое биологическое значение имеет эта форма сплайсинга.
альтернативный сплайсинг–форма сплайсинга при котором соединения экзонов в процессе созревания мРНК происходит в разных комбинациях. При этом порядок расположения экзонов не нарушается. Сплайсинг обеспечивает кодирование генов разных белков, что является механизмом белкового разнообразия у эукариот(более 70% генов человека подвергаются альтернативному сплайсингу).
Транскрипция - это синтез РНК на матрице ДНК. У прокариот синтез всех трех видов РНК катализируется одним сложным белковым комплексом - РНК-полимеразой.
Синтез мРНК начинается с обнаружения РНК-полимеразой особого участка в молекуле ДНК, который указывает место начала транскрипции - промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся, и на одной из них фермент осуществляет синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементарности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. РНК-полимераза способна собирать полинуклеотид лишь от 5"-конца к 3"-концу, матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та, которая обращена к ферменту своим 3"-концом (3" → 5"). Такую цепь называют кодогенной.
Терминатор - это участок, где прекращается дальнейший рост цепи РНК и происходит ее освобождение от матрицы ДНК. РНК-полимераза также отделяется от ДНК, которая восстанавливает свою двухцепочечную структуру.
Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор, образует единицу транскрипции - транскриптон.
Оперонная регуляция (т. е. регуляция на уровне транскрипции) – основной механизм регуляции активности генов у прокариот и бактериофагов.
Оперон - участок генетического материала, транскрипция которого осуществляется на одну молекулу иРНК под контролем белка-репрессора.
Оперон состоит из тесно сцепленных структурных генов, кодирующих белки (ферменты), осуществляющие последовательные этапы биосинтеза какого-либо метаболита. Каждый оперон содержит: промотор, оператор, и терминатор.
Оператор - нуклеотидная последовательность, связывающая репрессорный белок и негативно регулирующая транскрипцию соседнего гена . Оператор находится между промотором и структурными генами. Он может быть связан с особым белком - репрессором, который не дает двигаться РНК-полимеразе по цепи ДНК и препятствует синтезу ферментов. Таким образом, гены могут включаться и выключаться в зависимости от наличия в клетке соответствующих белков-репрессоров.
Репрессор - регуляторный белок, подавляющий транскрипцию генов регулируемого им оперона в результате связывания с оператором (регуляторным участком оперона). Это приводит к прекращению синтеза соответствующей иРНК и, следовательно, ферментов, кодируемых опероном. Репрессор синтезируется под контролем гена-регулятора в кол-ве от 10 до 20 молекул на клетку в виде активной, т. е. способной непосредственно связываться с оператором, или неактивной форм. Образование активного репрессора характерно для индуцибельных ферментов, синтез которых начинается только при попадании в клетку специфических низкомолекулярных веществ - индукторов. Индуктор - небольшая эффекторная молекула, связывающаяся с регуляторным белком, или физический фактор (свет, температура), которые стимулируют экспрессию генов, находящихся в неактивном состоянии.
Для осуществления правильной транскрипции необходимы регуляторные элементы двух типов. Регуляторные элементы первого типа называют цис-регуляторами . Они представляют собой специфические последовательности ДНК на данной хромосоме. Цис -регуляторы оказывают действие только на ближние гены. Второй тип называют транс-регуляторами. Это растворимые молекулы (включая белки и РНК), которые продуцируются одним геном, а взаимодействуют с другими генами на той же хромосоме или на других хромосомах. Если обратиться к индукции генов в lac -опероне Е. coli , то можно вспомнить, что ген репрессора дает белок-репрессор, который взаимодействует с последовательностью оператора для генов lac -оперона. В этом случае оператор является цис -регуляторным элементом, так как он контролирует только lac - оперон своей собственной хромосомы. (Последовательность мутантного оператора на другой хромосоме может присоединять или не присоединять белок-репрессор.) Белок-репрессор, напротив, является транс -регулятором. поскольку он продуцируется одной хромосомой, а связывается с цис -регуляторным оператором на другой хромосоме (рис. 12.5).
В эукариотических генах, кодирующих мРНК, обнаружены два типа цис -регуляторных последовательностей ДНК – промоторы и энхансеры («усилители»). Промоторы обычно располагаются непосредственно перед сайтом, в котором начинается
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
142 _______________ ГЛАВА 12 _____________________________________________________________________________
|
Рис. 12.5. Схема дифференциальной регуляции гена у E . coli ; показаны цис - и транс -регуляторные элементы. В клетках дикого типа индуцибельное состояние характеризуется тем, что РНК для β-галактозидазы не транскрибируется, пока отсутствует лактоза. В отсутствие лактозы белок-репрессор (R) кодируемый геном i , присоединяется к сайту оператора (о ), ингибируя этим транскрипцию РНК-полимеразой с промотора (p ). Если лактоза присутствует, то она связывается с белком-репрессором, в результате репрессор не может присоединиться к ДНК и транскрипция продолжается. Растворимая природа этого репрессора показана в опытах на мутантах E . Coli . Когда гаплоидные бактериальные клетки, несущие ген i – , становятся частично диплоидными с геном i дикого типа (i + ), синтезируется репрессор дикого типа, который способен сделать индуцибельным исходный ген ß-галактозидазы. Этот белок-репрессор является транс -регуляторным элементом. Последовательности промотора и оператора представляют собой цис -регуляторные элементы. |
|
|
|
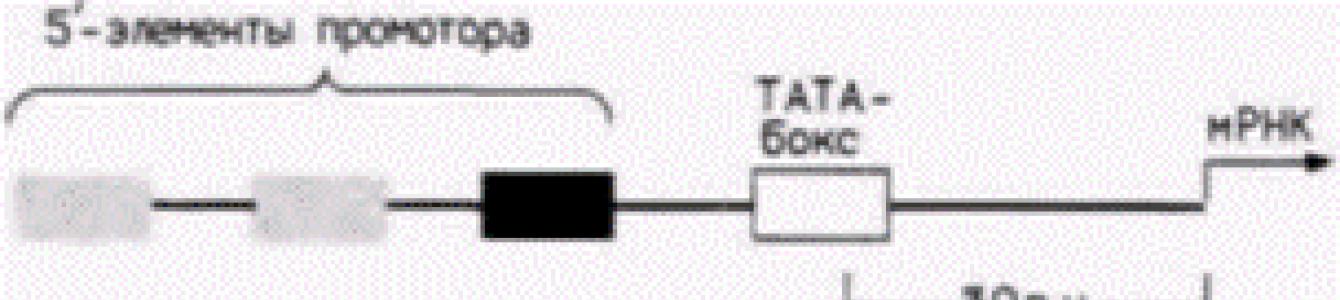
Рис. 12.6. Типичный промотор для гена эукариот, кодирующего белок. Представленный ген содержит ТАТА-бокс и три 5"-элемента промотора. Примеры таких 5’-элементов представлены в нижней части рисунка. (По Maniatis et al., 1987.) |

транскрипция, и длина их составляет приблизительно 100 пар оснований. Участок промотора необходим для присоединения РНК-полимеразы II и точной инициации транскрипции. Энхансер активирует утилизацию промотора, контролируя эффективность и скорость транскрипции с этого конкретного промотора. Энхансеры активируют только лежащие в цис -положении промоторы (т.е. промоторы на той же самой хромосоме), но они могут функционировать и на больших расстояниях. Кроме того, они могут находиться не только на 5"-стороне гена, но и на другой цепи ДНК (Maniatis et al., 1987).
Промоторы генов, которые транскрибируют относительно большие количества мРНК, имеют сходную структуру. В них содержится последовательность АΤΑ (называемая иногда ТАТА-боксом или боксом Голдберга–Хогнесса ), располагающаяся на расстоянии приблизительно 30 пар оснований с 5"-стороны от сайта, где начинается транскрипция, и один или несколько передних элементов промотора , лежащих еще дальше с 5"-стороны. Передний элемент промотора обычно представляет собой вариацию последовательности ЦААТ, но выявлены и другие промоторные элементы (Grosschedl, Birnstiel, 1980; McKnight, Tjian, 1986) (рис. 12.6).
Впервые промотор β-глобинового гена исследовали в опытах по проверке специфической транскрипции клонированной ДНК. Клонированные гены могут транскрибироваться правильно, когда они введены в ядра ооцитов лягушки или фибробластов или когда они инкубируются с очищенной РНК-полимеразой в присутствии нуклеотидов надосадочной жидкости (Wasylyk et al., 1980). После того как транскрипция гена подтверждена, для получения специфических делений в этом гене или окружающих его участках используют рестриктазы. Затем можно выяснить, продолжает ли правильно транскрибироваться такой модифицированный ген. Результаты этих исследований показали, что для максимальной транскрипции ß-глобинового гена достаточно первых 109 пар оснований, предшествующих кэп-сайту (Grosveld et al., 1982; Dierks et al., 1983).
Другие исследователи уточнили этот вывод с помощью клонирования участка глобинового гена мыши от 106-й пары оснований выше (с 5"-стороны) старта транскрипции (положение -106) вплоть до 475-й пары оснований (положение +475) в первом экзоне (Myers et al., 1986). Эти клоны были подвергнуты мутагенезу in vitro. Таким способом в область промотора глобинового гена было введено






